Unique features of a global human ectoparasite identified through sequencing of the bed bug genome
摘要:温带臭虫Cimex lectularius,在过去的二十年里,是世界范围内广泛存在的一种人体皮外寄生虫。除了对杀虫剂抗性的增强,这种全球化范围的害虫增加可能与国际交流和贸易有关。温带臭虫基因组(650Mb)及其14,220个编码蛋白质的基因全面展示了其与创伤性受精(traumatic insemination)、减少化学感应相关的专性吸血基因、宿主-共生体的相互作用以及几个对杀虫剂抗性的机制相关的基因。基因组测序和注释为未来对温带臭虫杀虫剂抗性、人类-臭虫和共生体-臭虫的相互作用关系、臭虫作为人类皮外寄生虫独特的生物学特性提供了坚实的基础。
温带臭虫Cimex lectularius,证明与人类有3000年甚至更多年的联系。二战后,由于杀虫剂的自由使用,这个物种在一些经济和政治稳定的国际几乎灭绝,但是在一些不发达国家、弱势地区还有大量的与老鼠、鸡和其它动物相关的群体存在。然而,在过去的20年左右,温带臭虫在除南极洲外的每个大陆大量的爆发。温带臭虫的快速扩张与国际旅行、二手材料的转换、对有温带臭虫组织的无知、臭虫对有机氯、有机磷酸酯类和拟除菊酯等主要杀虫剂的抗性进化。
作为人类皮外寄生虫,温带臭虫的成功扩张是由其以下几个方面的生物特性造成的。首先,温带臭虫的所有生命阶段都专性地食血,并以血液作为单一的营养和水分来源。这种营养的专一化需要专性的化学感应系统来探测、发现和接受合适的宿主。并且,两个不同的温带臭虫家系研究表明,与人类有关和与老鼠有关的两个家系有不同的化学感应特化。另外,吸血作用需要特殊的酶和相关的代谢通路来适当地消化和吸收血液,并处理多余的水分;这种专化作用也驱动与内共生体的转化关系,例如沃尔巴克氏体 (一种革兰氏阴性细菌)可以产生脊椎动物血液中缺乏的关键微量元素。
温带臭虫通过创伤性受精来繁殖,雄性用改进的生殖器官刺进雌性腹部的角质层,将精液带入雌性的淋巴系统。这种繁殖方法受到性别冲突的强烈选择,包括性传播细菌,及选择免疫系统来影响臭虫-病原菌的互作。臭虫表现出对近亲繁殖作用的抗性,这可能与有益基因复合物的固定,促进局部适应的有毒等位基因,如对杀虫剂抗性基因的清除有关。有意思的是,远亲繁殖没有表现出劣势,而是杂种优势最小化且短暂。温带臭虫被认为是美国锥虫的潜在载体,不像其它的吸血节肢动物,可以直接确定人类病原物的传播。最后,具有杀虫剂抗性的种群大量扩增,具有拟除菊酯抗性的温带臭虫数量是没有抗性的10,000倍。
通过进化基因组分析和大量的人工注释,本文报道了温带臭虫极其相关的细菌基因组。本研究揭示了温带臭虫与其生活方式相关的进化适应,包括化学感应基因的显著减少,血液代谢相关基因的扩张和各种其它物种与杀虫剂抗性相关的基因集。另外,研究人员鉴定了从沃尔巴克氏体和沙雄菌等细菌中横向迁移(lateral gene transfer,LGT)的基因。
对各种杀虫剂敏感的温带臭虫,进行基因组测序,并作为转录组测序中的对照处理。在27 ℃, 50±5% RH和光周期12:12(L:D)条件下保存培养,喂食37℃水域保温的兔子血。取第23代自交系的个体基因组DNA。
Illumina HiSeq2000s测序平台,PE100。小片段文库180bp,500bp,大片段文库3kb,8kb,测序深度分别为62.4X, 77.9X, 44.42X, 21.21X。
1基因组的总体特性和直系同源分析
基因组最终组装成650.47Mb,1,402 个scaffolds和45,073个 contigs,N50 长度分别为7.17Mb and 23.5 kb。比预估的基因组大小(864.5Mb)小25%,这可能是由于异染色质和重复序列没有组装的结果。预测了14,220个蛋白质编码基因,总共有1,352个是感兴趣的基因,包括273个角质层蛋白基因和114个化学感受体基因进行了人工校正。评估了组装和基因预测的完整性,鉴定了昆虫基因组中保守的Hox基因簇和自噬基因。总的来说,与七种节肢动物昆虫基因组相比,温带臭虫基因集和基因组有少量的的丢失,大约10%,但是仍然具有很高的完整性,来进行进一步的分析。温带臭虫中,超过80%的基因至少与一种节肢动物具有直系同源基因。用8个节肢动物的1,734个单拷贝的直系同源基因来构建系统进化树。

图1 温带臭虫和其它节肢动物的系统进化关系及直系同源比较
注:系统进化分析表明温带臭虫是另一种半翅目昆虫-豌豆蚜的姊妹系。
2宿主定位、专性血液为食和免疫
昆虫感受器中嗅觉和味觉的处理是依靠化学感受体的三个基因家族:嗅觉受体、味觉受体和离子受体。嗅觉受体在黑暗环境中通过嗅觉来寻找同伴(mate)、宿主定位(host location)和导航中起到关键的作用。味觉受体(GRs)的主要作用是调解味觉—更主要的是发现甜(吞噬刺激)或苦(威慑)的增味剂—以及感受二氧化碳。亲离子受体由古动物的离子移变的谷氨酸盐受体进化而来,与嗅觉和味觉有关。作者鉴定了48个嗅觉受体、24个GRs、30个亲离子受体基因。温带臭虫化学感受体基因集居中,与其适中的化学生态的复杂性相一致,它像采采蝇(Glossina morsitans)一样是专性的吸血昆虫,但是具有广泛的脊椎动物寄主,而虱(Pediculus humanus)则以人类作为唯一的寄主。臭虫基因组中没有糖受体,而在采采蝇、虱子等吸血性昆虫当中也没有发现。臭虫基因组中化学感受体基因家族有一些扩张和进化,同时具有11个物种特异性的增味剂结合蛋白,14个化学感受体蛋白比其它吸血性昆虫具有较强的保守性。
获得血液的一个主要障碍是宿主止血,宿主通过血小板聚集、纤维蛋白交联、血管收缩和免疫应答来防止血液的流失。臭虫唾液转录组和蛋白质组的建立,有助于我们对其唾液的复杂性、吸血昆虫独特的适应性进行研究。臭虫三磷酸腺苷双磷酸酶有多个成员,参与依赖ADP的血小板聚集抑制蛋白。另外,鉴定了12个一氧化氮受体多磷酸肌醇磷酸酶基因家族成员,6个昆虫基因组中最大的水解酶家族Ap4a家族成员。唾液蛋白的扩增使臭虫能够从同一宿主多次取食血液,而不会使宿主痛苦。
脊椎动物的血液富含蛋白质和脂类,但是缺少微量营养。由于其较高的水分含量,需要特定的一套消化酶来消化。对臭虫基因组的分析,发现了187个潜在的消化酶基因。与大多数昆虫相比,臭虫具有较少的丝氨酸蛋白酶,但是其蛋白酶与吸血的长红猎蝽、虱相似。臭虫的组蛋白酶D大量扩张,它是一种调节酸性pH的天门冬氨酸蛋白酶,被认为与血液的代谢有关。
血液的消化需要将其中多余的水分移除,水通道蛋白(AQPs)在这一过程起重要作用。臭虫有7或8个AQP基因,在大多数昆虫中6-8个为正常范围。在肽类激素和胺受体中,作者鉴定了一个完整的利尿和抗利尿激素及其受体精确地启动和终止利尿。与长红猎蝽不同,臭虫质有一个基因编码抗利尿神经肽激素。
像吸血蜱和锥蝽,但不像其他吸血昆虫,臭虫可以经受长时间的饥饿。这种适应需要营养的保持(例如较低的代谢)和阻止多余水分散失、脱水导致的死亡,后者尤其依赖于水通道蛋白和热激蛋白的差异表达。总的来说,臭虫中热激蛋白和自噬基因与其它昆虫类似,这表面你给它们的不同可能是与极端脱水和抗饥饿有关。这些基因序列可以帮助人们揭示臭虫耐极端脱水和饥饿的生理和行为机理。
臭虫的基因组中具有较多的Toll,Imd和Jak/STAT免疫通路相关的基因。

图2 臭虫基因组宿主定位(host location)和吸血方面相关的鉴定
a:多种昆虫的化学受体相关基因;b:多种昆虫的唾液功能相关基因;c:多种昆虫组蛋白酶D构建的系统进化树。
3共生和横向基因迁移
由于血液中营养物质的不足。专性吸血作用会导致特殊微量元素的缺乏。沃尔巴克氏体是昆虫中常见的一种体内寄生虫,可以影响寄主的生长和繁殖,它与臭虫进化出了共生的营养关系。沃尔巴克氏体为臭虫提供繁殖和发育所需的各种维生素B。作者注释了维生素B代谢和决定相关的基因,与其它昆虫类似,臭虫中含有维生素B摄取和转换(salvage and conversion)必需的基因。
用计算管道从组装好的臭虫基因组中检测细菌基因组,作为候选的细菌中横向迁移的基因。温带臭虫的内共生体沃尔巴克氏体组装成16个scaffolds。另外,与温带臭虫相关的葡萄球菌基因组几乎组装完整,包括15个scaffolds,其中有3个质粒和一个大约3.16Mb的染色体。葡萄球菌在臭虫中体内常见,并且通过性接触传播。第三个sacfford与果胶杆菌具有同源序列组装成,这种菌与植物相关,sacfford 只有250kb大小,而果胶杆菌基因组大约有5Mb,作者推测它是臭虫中大片段的横向基因插入引起的,还需要进一步的研究来确认。
臭虫基因组中有大量的细菌LGT(横向迁移)基因。除了以上讨论的部分,有805个>100 bp候选LGTs散在分布在整个基因组上。 臭虫基因组中候选LGTs在14种节肢动物中数量最多。来自杀雄菌属的LGTs数量最多,在已经测定基因组的昆虫中非常罕见。臭虫中的LGT是独特的插入序列,而在其它的昆虫基因组中则没有发现。
LGT基因插入后,一般通过插入(由于非同源DNA修复机制)及降解和丢失来进化。因此,LGTs与宿主DNA结构类似。然而,有些真菌LGTs可以进化出具有功能的基因。臭虫中大多数的候选LGTs在成熟的个体中不表达,有一个例外,臭虫中的来源于沃尔巴克氏体LGT基因,编码马铃薯样蛋白的基因。细菌中马铃薯样蛋白基因与其致病性相关。臭虫中的这个LGT在雄性成熟个体中表达,而在雌性成熟个体中不表达。这一基因的功能还需要进一步确定,但是可以推测它与臭虫特殊的受精过程有关。
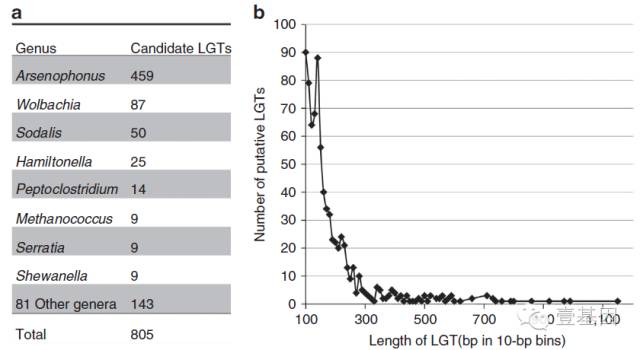
图3 臭虫基因组组装后>100 bp的候选横向基因迁移(LGTs)
a:鉴定的候选LGTs数量;b:候选LGTs的长度。
4杀虫剂抗性相关基因
过去的二十年,臭虫对拟除菊酯抗性的增强导致其大面积爆发。为了分析其原因,作者对有杀虫剂抗性的昆虫进行了转录组测序,分析了与杀虫剂抗性相关的基因。电压门控钠离子通道基因的两个突变体可以使臭虫具有拟除菊酯抗性。细胞素色P450s在臭虫对杀虫剂的抗性中也起作用。ATP-结合盒(ABC)转运体与外源物质的转运有关,在具有杀虫剂抗性的昆虫中量增加,臭虫中一共有54个ABC转运体。羧酸酯酶在杀虫剂的代谢中起到关键的作用,具有杀虫剂抗性的臭虫中,编码这个酶的基因大量复制。角质层在臭虫对杀虫剂的抗性中从很多个方面起作用,鉴定了273个角质层基因,其中CPR基因家族出现扩增。
5创伤性受精
臭虫的雌性进化出了一个新的器官,可以通过超弹性节肢蛋白来减少交配时产生的创伤。作者对基因组的分析,发现臭虫节肢弹性蛋白基因家族扩张,有13个基因具有几丁质结构域,并富含>20%的甘氨酸。这个基因家族的扩张是臭虫特有的。
臭虫基因组的序列将帮助人们了解它作为人体皮外寄生虫的生物学特性,如图4。
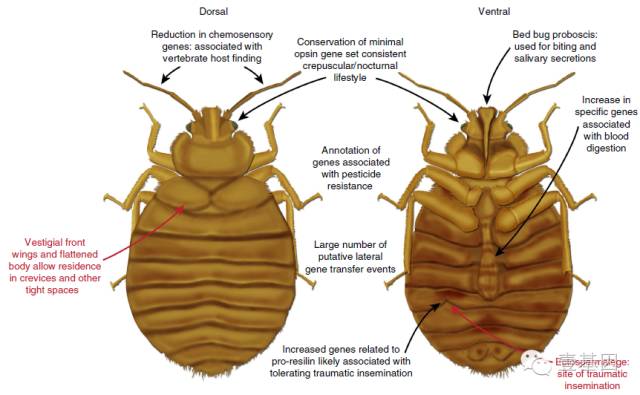
图4 臭虫基因组关键生物学过程概要
注:红色:臭虫的一般生物学特性;黑色,通过基因组序列分析和人工校正获得的关键生物学问题